Meet your colleagues and discuss a new preprint over lunch during #cellbio2023 🥳
We’re pleased to announce preprint launch parties organized by ASAPbio on Dec 4 & 5 in Boston, MA, USA!
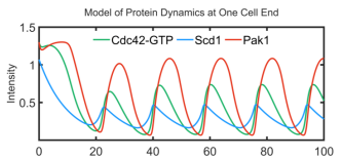
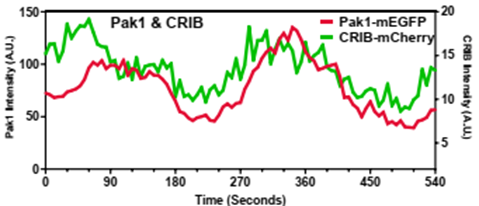
Our models suggest that there is a phase shift between the accumulation of Cdc42, Scd1, and Scd2 (Positive feedback loop) and Pak1 (time-delayed negative feedback). Indeed, we were able to capture these shifts in vivo. (Colors correspond to protein of interest) (12/17)
I am pleased to announce that my first manuscript as fist author is now posted on @biorxiv_cellbio. Thank you to Ziyi Liu (co-first author), @bethfcampbell1, Livi Chinsen, @hongtian162 and especially to my graduate mentor Dr. Maitreyi Das @DasLab_Pombe (1/17)
And while much work has established the impact of Cdc42 on membrane trafficking, we are now showing that membrane trafficking, in turn, directly regulates Cdc42 activity. (16/17)
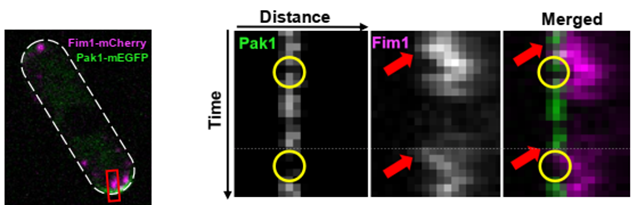
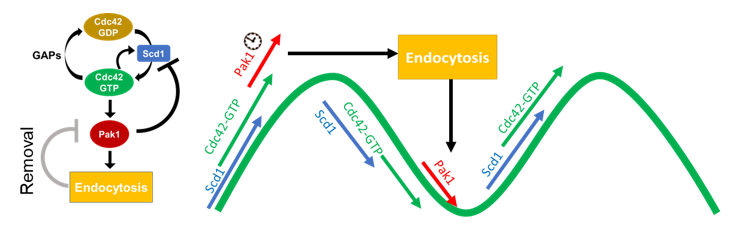
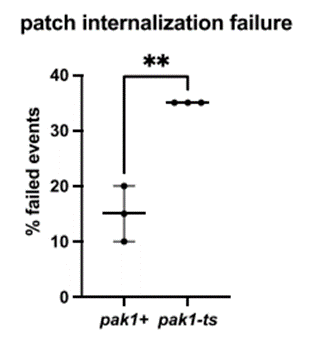
Further, we show the regulatory relationship between Pak1 and endocytosis. We find that Pak1 is removed from the membrane as endocytic vesicles internalize and, simultaneously, Pak1 activity is required for proper vesicle internalization. (11/17)
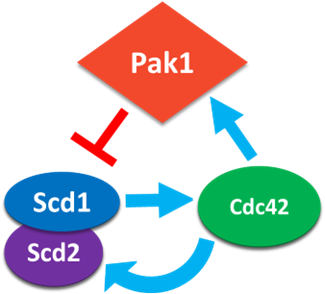
Anticorrelated oscillations are the result of positive feedback (Scd1 & Scd2) and time-delayed negative feedback (via the Pak1 kinase). The anticorrelated patterning also suggests that the ends must compete for Cdc42 activity. (5/17)
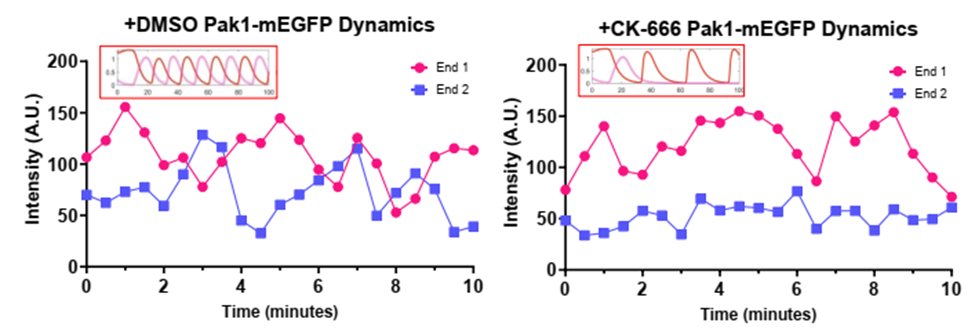
We find that endocytosis is required for proper Cdc42 activity between ends. Using in vivo experiments alongside mathematical modeling, we show that Pak1 stabilizes at a cell end when endocytosis is disrupted, preventing the return of Scd1. (Model in insets) (10/17)
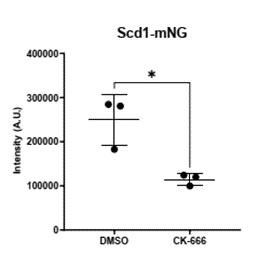
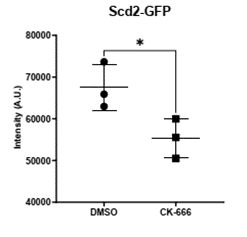
Normally, Arp2/3 is required for endocytosis to recycle protein from the membrane in yeast. But, surprisingly, localization of the Cdc42 activator Scd1 and its scaffold Scd2 are significantly decreased when the Arp2/3 complex is inhibited. (9/17)
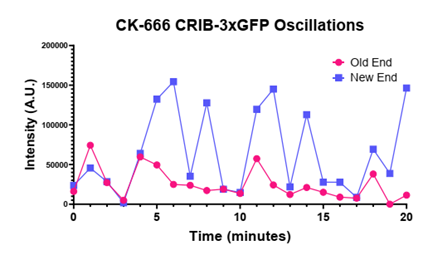
Unexpectedly, we find that cells are no longer able to reestablish Cdc42 activity at a second end when the Arp2/3 complex and branched actin were disrupted. (8/17)
we observed that F-actin structures (branched actin and actin cables) are required for oscillations. So, we asked is competition for Cdc42 wired or wireless? Is it dependent on cables (which span the cell like phone lines) or branched actin (which don’t contact both ends)? (7/17)