Postdoc at David Baker lab @ IPD U Washington | PhD at Cees Dekker lab @ TU Delft | Schmidt Science Fellow | Creating and understanding the tiniest machines
Exciting news 📃🧬🔬 Today we report on the dynamics of a fascinating class of molecular motors, so-called SMC proteins in @CellCellPress
https://t.co/MRIIHE893Z
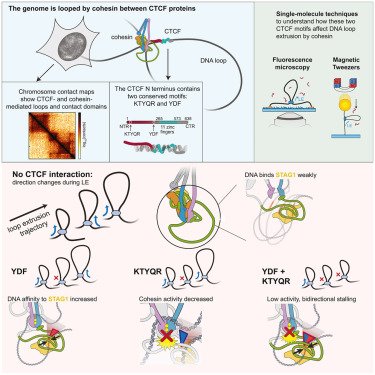
🚨New paper out! In our @MolCellNews paper, we solve a piece of the puzzle of how our genome folds. We found the two specific molecular "brakes" in the CTCF protein that stop DNA loop extrusion. One of the final pieces of my PhD!
https://t.co/SYa5Y2ZeQT
CTCF doesn't just have one stop signal—it has two!The YDF & KTYQR motifs stall the cohesin loop extruder in different ways: one is a roadblock, the other forces one-way traffic.
A huge congratulations to the recipients of the 2025 Weintraub Award! These ten exceptional graduate students exemplify the bold, creative, and pioneering spirit embodied by Dr. Hal Weintraub. https://t.co/DqyAYoQ2ki
I'm excited to share that I've been selected as one of the 10 recipients of the prestigious Weintraub Award from
@fredhutch this year! 🏆 I owe a massive debt of gratitude to the remarkable colleagues and mentors who have shaped my path, especially @cees_dekker
!!
Science is a collaborative adventure, and this moment really feels like a celebration of the collective effort, curiosity, and passion that drives research.
@TNWTUDelft
Thank you to all that contributed!! Especially to @cees_dekker and the fantastic team at the @IMPvienna Gabriele Litos, Iain Davidson, and Jan-Michael Peters!!
KTYQR is distinct from YxF: it reduces cohesin's LE activity, but doesn't impact its directionality. Together, the two account for most of the impact of the full NTR! We hypothesize that connecting the two on one protein chain and some adjacent sequences modulate further.
We hope that by disentangling the multiple contributions of the CTCF N-terminus to the stalling of cohesin at CTCF sites, we are another step closer to understanding how cohesin and CTCF control and shape genomes across the tree of life 🌳
Even though YxF has a large impact on cohesin's LE activity, it doesn't fully account for the impact of the full N-terminal region of CTCF. So we searched more 🔎
Curiously, we observed that cohesin also changes direction much less frequently in the presence of YxF! With inspiration from Shaltiel et al, 2022, we hypothesized that this is due to a higher DNA affinity of STAG1-kleisin in the presence of YxF
CTCF stall DNA 🧬 loop ➰ extrusion by cohesin - but exactly how it pulls this off is a 'mechanistic mystery' (to cite
@Anders_S_Hansen , Nucleus, 2020). We just preprinted 📜 a new study @biorxivpreprint that provides some answers to this enigma: https://t.co/CYYDP0NvrE
We started with the known YxF motif. In brief, the YxF motif:
- reduces cohesin's ATPase rate
- reduces LE initiation
- reduces the fraction of complete LE steps (in Magnetic Tweezers)
So these few amino acids alone already tinker quite a lot with cohesin!
Observing LE by Magnetic Tweezers provides a much more detailed view. We can see individual steps! Previously, we already observed a plethora of phenomena: cohesin often makes full LE steps, as seen by a height decrease of the magnetic bead. But these are also often reversed
Single-molecule fluorescence visualization lets us assess quantitative metrics such as the loop extrusion (LE) initiation rate, LE rate, directionality changes (see https://t.co/dciPuqjDQ4 ..., etc.
We took the CTCF N-terminal region that is already well-known to interact with cohesin and cut it up into smaller pieces. Then, we exposed cohesin to these various fragments in buffer (no DNA binding domain here!) and observed how cohesin handles that.