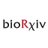1/ Thrilled to share our new paper in @Nature: “CLCC1 promotes hepatic neutral lipid flux and nuclear pore complex assembly.” From our lab together with @apsarruda, led by co–first authors @alymathiowetz and @EmilyMeymand. 🥳
https://t.co/LHO31rNaxi
Also, quite coincidentally to me working at @UCLeye, a mutation in CLCC1 causes a rare autosomal recessive #Retinitis (RP32)
https://t.co/fEAKCJelka
Is the molecular basis of that blinding disorder related to nuclear pore complex insertion? Or another nuclear envelope function?
Great to see our preprint out. We uncovered a simple likely mechanism to fuse inner+outer nuclear envelopes to precede nuclear pore complex (NPC) insertion.
Our first step was to show that CLCC1 is the long-lost human homologue of Brl1 (previously only known in yeast, protists)
3/3 A major feature of these rings is that they deform the membrane into the lumen. Two opposing rings on inner and outer nuclear envelopes might lead to fusion, forming a pore that then opens to accommodate the NPC.
Congratulations to @alymathiowetz & the whole team.
Great to see our preprint out. We uncovered a simple likely mechanism to fuse inner+outer nuclear envelopes to precede nuclear pore complex (NPC) insertion.
Our first step was to show that CLCC1 is the long-lost human homologue of Brl1 (previously only known in yeast, protists)
New preprint w/ @apsarruda! Tour de force from grad student extraordinaire @alymathiowetz! >20 CRISPR screens enabled the discovery of a key role for CLCC1 in hepatic neutral lipid flux and nuclear pore complex assembly. Many surprises! #LipidDroplets 1/3 https://t.co/Z5iJioxk1l
2/3
Interestingly both CLCC1 (human) and Brl1/Brr6 (yeast) both have strong lipid droplet phenotypes too.
Next we made AF2 predictions of oligomers. Both proteins are predicted to form ring oligomers.
Here's the CLCC1 ring (yellow=TMH, pink=amphipathic, same as Brl1 ring above)
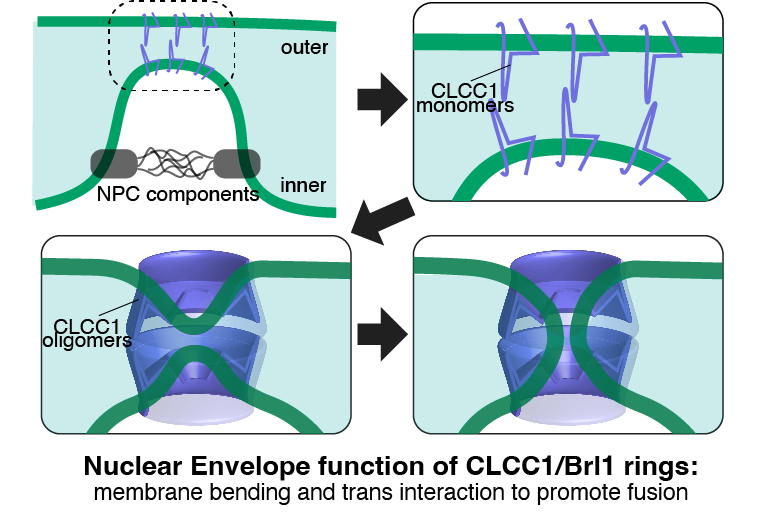
Yaoyang Zhong, PhD student in my lab, & I have a new News and Views in @ContactCTC about the discovery by Karin Reinisch's team (Yale) that the senescence domain in Spartin is a lipid transfer protein https://t.co/rZYRv2NiWR
The original work is in PNAS: https://t.co/609U3yFYIt
Excellent new paper from the lab of Sabrina Buttner (& collaborators incl. me!) showing LDO proteins bridge from lipid droplets to the yeast vacuole to allow lipophagy during starvation.
Could the animal LDO-ologue promethin (=LDAF) act similarly? it has ~30aa disordered tail.
We are so proud to share the final version of our latest #MembraneContactSites story🎉, now in @Dev_Cell. We identify the LDO proteins and Vac8 as a tethering pair between the vacuole and lipid droplets (vCLIP) that mediates microlipophagy in starvation
https://t.co/jHAhSJWvVf
Our lab has a new preprint out on how LAM proteins self-organise into puncta. This turns out to NOT positively affect function ... rather the opposite.
https://t.co/DhC9Tnml6J
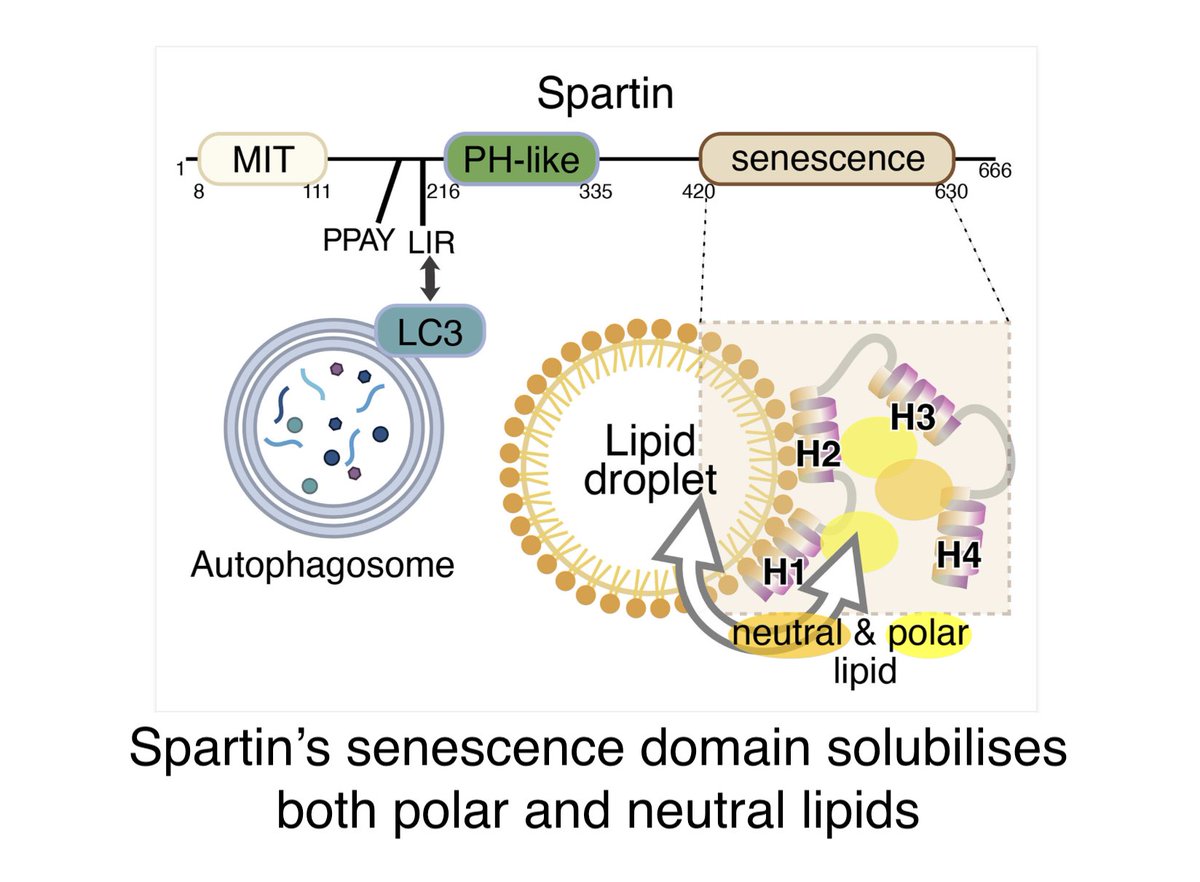
+We collaborated with @KukulskiWanda to idenitfy the ER architecture at LAM+ puncta: it's unremakrable.
Elife is moving forward but its claim to be “relinquishing role of gatekeeper” applies only to removing gate #2.
Elife will still keep gate #1: “we do not have the capacity to review every preprint submitted to us”. Deciding what to review is now uniquely important... & secret!
Massive news: eLife to abolish accept/reject decisions: papers will just be “peer reviewed”. Others can argue about this, but lots of interesting consequences. 1/9 https://t.co/FedRxI62iC
5 of 5
Most RBG multimers have two amphipathic helices that cross over the groove at their N- and C- ends. SHIP164 is the exception, with its C-terminus lacking this kind of helix, but having a coiled coil. The paper speculates on what role that might play.
My paper about VPS13 ATG2 +3 other eukaryotic bridge-like lipid transfer protein families made from repeating beta groove (RBG) domains is now out in
@ContactCTC .
https://t.co/C9TsYEej9A
I uses sequence homology & structural predictions to analyse the proteins
(1 of 5)
4 of 5
Viewing the most conserved residues in detail shows that there are many hydrophilic residues all the way along the outside of the lipid-binding groove, well placed to bind protein partners.
3 of 5
Most RBG domains are highly conserved across all orthologues, and many are conserved between RBG proteins.
However, some are highly variable indicating that sequence conservation is required for a function other than lipid transfer, likely to be interacting with partners.
2 of 5
Comparing all the RBG domains in VPS13 with each other identified just four types:
• the extreme ends (x2)
• just 2 central types (here shown in blue/red) that have duplicated many times
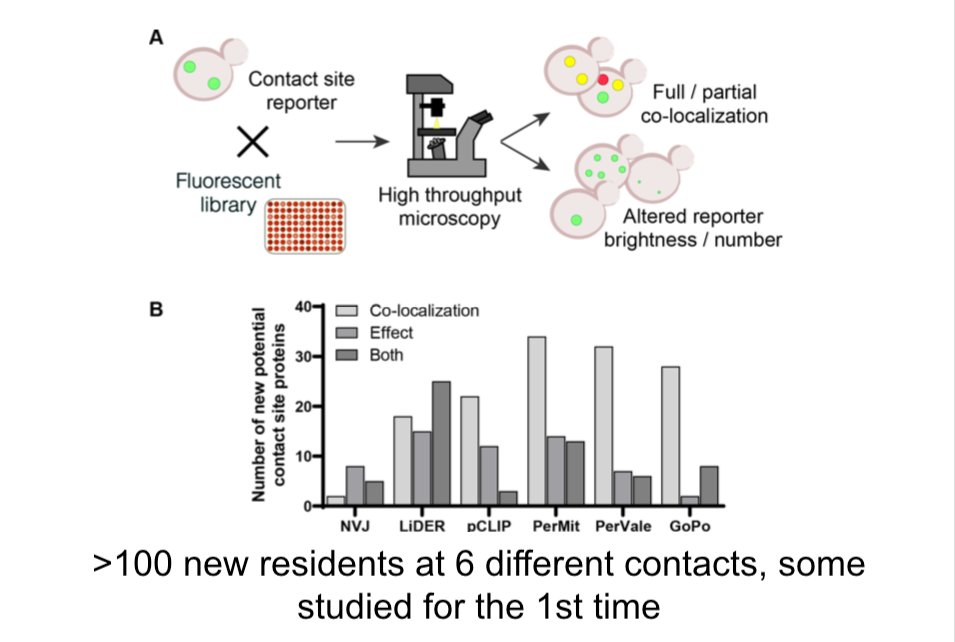
Our 6-way collaboration is out, led by Schuldiner lab @WeizmannScience, @conibear_liz lab @UBC & Chris Stefan @LMCB_UCL. Also Vanina Zaremberg @BioSciUofc & Robert Ernst SaarlandU
We found lots of new contact site proteins, many of which transfer lipid
https://t.co/4a7uhF9d2l
For some years we've known that lipids flow between organelles on static bridges (VPS13, ATG2 …)
AlphaFold allowed us to define a Repeating Beta Groove (RBG) domain in all the bridges https://t.co/uf74TjL2pg
Now here's new detail showing how RBG superfamily bridges might work
Sequence analysis and structural predictions of lipid transfer bridges in the repeating beta groove (RBG) superfamily reveals past and present domain variations affecting form, function and interactions of VPS13, ... https://t.co/0137sLU1BP #bioRxiv