Temporarily reactivating regenerative genes can heal injuries and cure degenerative diseases of old age by unlocking regenerative capacity from early life.
Continuing with part 3/10 of the trail that led to developmental age reversal, I began my doctoral research in 1982 with Samuel Goldstein, MD, a brilliant aging researcher being the first to use the then emerging field of recombinant DNA research to study aging using the Hayflick model of fibroblast senescence. Sam had an uncanny instinct that it was changes in repetitive DNA sequences or DNA methylation that was a root cause. Unfortunately, I ended up disproving his published results on the loss of EcoR1 repeats and Inter-Alu (the latter being a provocative paper published in Nature and subsequently retracted).
Not in alignment with Sam on how to handle this conflict, I decided to start my PhD afresh at Baylor College of Medicine and wrote a dissertation on my discovery that senescent cells secreted destructive proteolytic enzymes that tear down tissues faster than they could be regenerated (what Judy Campisi later called the Senescence-Associated Secretory Phenotype (SASP)).
Then later as a medical student at UTSW in Dallas, I stumbled on a paper by a 1986 paper published by Howard Cooke titled “Variability at the Telomeres of the Human X/Y Pseudoautosomal Region” where he showed the age-dependent shortening of telomeres in somatic cells but maintenance at a long length in germ line cells. This fit the Weismann model exactly. This, together with work I was doing with Woody Wright on cells in extreme senescence (Mortality 2) where chromosomes began joining end-on-end, the data convinced me that a wild-eyed theoretical scientist named Alexey Olovnikov was right. The clock of cell aging was the loss of DNA at the ends of the chromosomes and an unidentified protein then called “telomere terminal transferase” (now called “telomerase” for short), and the basis of cell immortality. So, Sam Goldstein instinct was right, I surmised, it was repetitive DNA loss, just not the sequences he was studying. Below is a video where I asked Alexey in St Petersburg to recount how he dreamed up the hypothesis. This theory turned out to be true as we demonstrated once we had found the telomerase gene as I will relate in the next post.
https://t.co/I8R5HcupRj
I was asked to clarify the history of the first persons’ cells to be immortalized by telomerase. Here’s what happened… About the time we had finally found the telomerase gene, Len Hayflick was visiting the lab and gave a film crew an illustration of how to culture skin cells by hoisting up a pant leg, grabbing a scalpel, and slicing off a piece of skin from his leg! Later he walked into my office and asked, “you want a piece of me?” I said, “yeah, let’s measure the true Hayflick Limit (how many doublings his cells would go before becoming senescent. I cultured his cells and was feeding them at the end of a days work and thought, history should have it that the first cells ever immortalized by human intervention should be Len’s own skin cells. So I took the newly-isolated telomerase gene that by then had been placed into a viral vector, and squirted it into the dish with his cells. I remember in my notebook I titled the experiment “The Immortalization of Dr Hayflick.” Weeks later when it became clear that it had worked, I had dinner with Len and his wife Ruth in a restaurant in Sonoma County on the sea coast. “I said, Len, about that experiment…” He said, “Yeah, what happened with that?” I explained how his untreated cells had senesced but his treated cells were growing exponentially and had clearly immortalized.” He stared out the window for the longest time then whispered “I can’t be that simple!” I tell the story because it’s a good example of how at first glance we think aging must be wear and tear and be very complicated (Len clearly thought that way). But the reality is that a single letter in the DNA code can cause premature aging in children with progeria, and a single gene (telomerase) can rescue cells from senescence. In upcoming posts I will explain how we figured out the other pieces of the aging puzzle, and like telomerase, it is far simpler that most people would have imagined.
In the previous post I discussed how August Weismann proposed that we are all are derived from a an immortal and regenerative lineage of cells called the “germ line.” He predicted that this amazing potential was lost when are bodies are being formed in the womb (something I call “somatic restriction”). Unfortunately, he was too far ahead of his time. About two years before Weismann’s death, Alexis Carrel (the first American to win the Nobel Prize authored a paper titled "On the Permanent Life of Tissues outside of the Organism.” Carrel’s paper convinced the scientific community that however we age, it was not an intrinsic clock within our cells. Charles Lindbergh, the famed aviator claimed that in his first trans-Atlantic flight, invisible beings in the back of his plane told him his destiny was to engineer human immortality. He sought out Carrel and together their theories on aging held sway until the 1960s when Leonard Hayflick overturned the prevailing dogma by demonstrating that body cells actually do age and that cell aging in the dish could serve as a lab model for deciphering the clockwork mechanisms of aging. That is the era I entered the field in search for the key to cell immortality. This was a first step toward human age reversal. I describe some of the drama of those early years in the YouTube video linked below.
https://t.co/ropJKC3Hh9
In this, the first of 10 posts on age reversal by genomic reprogramming, I begin where the story really needs to begin; namely, the insights of the German naturalist August Weismann. Weismann proposed the term “immortality” to describe the unceasing lineage of cells called the “germ line” that perpetuates the species (that is sperm and egg that make babies that make sperm and egg that make babies, ad infinitum…). Weismann theorized that cells that make up the body (somatic cells) lost that capacity for immortal regeneration, a phenomenon I call “somatic restriction.” He stated “Death takes place because a worn-out tissue cannot forever renew itself, and because a capacity for increase by means of cell division is not everlasting, but finite [in the soma].” If you think about it, this means that the cells that we are made of (trillions of them) have no dead ancestors. Tracing them back in time, the ancestors of the cells that make us “us” have been proliferating since the first dawn of life on Earth, some billions of years ago. They will abruptly die in a few decades in us. The question we began to ask nearly a century after Weismann was, “can we transfer the capacity of immortal renewal characteristic of the germ line cells into our body, into the soma.” In the next post I will describe the first of such steps that led to the modern technology of immortality transfer.
Over the next week I will share in a series of posts some insights and data on the reversal of human aging by transcriptional reprogramming (aka “partial reprogramming,” “epigenetic reprogramming of aging,” or “induced Tissue Regeneration (iTR).” Some of the critical aspects of the technology that need to be addressed are the fine tuning of the process to insure cells in a patient are adequately but not overly age-reversed, how telomere length (i.e. cellular age) can also be reversed, and how to prevent the current plurality of off-target effects of the first generation technology that carry potential risks of cancer. Stay tuned…
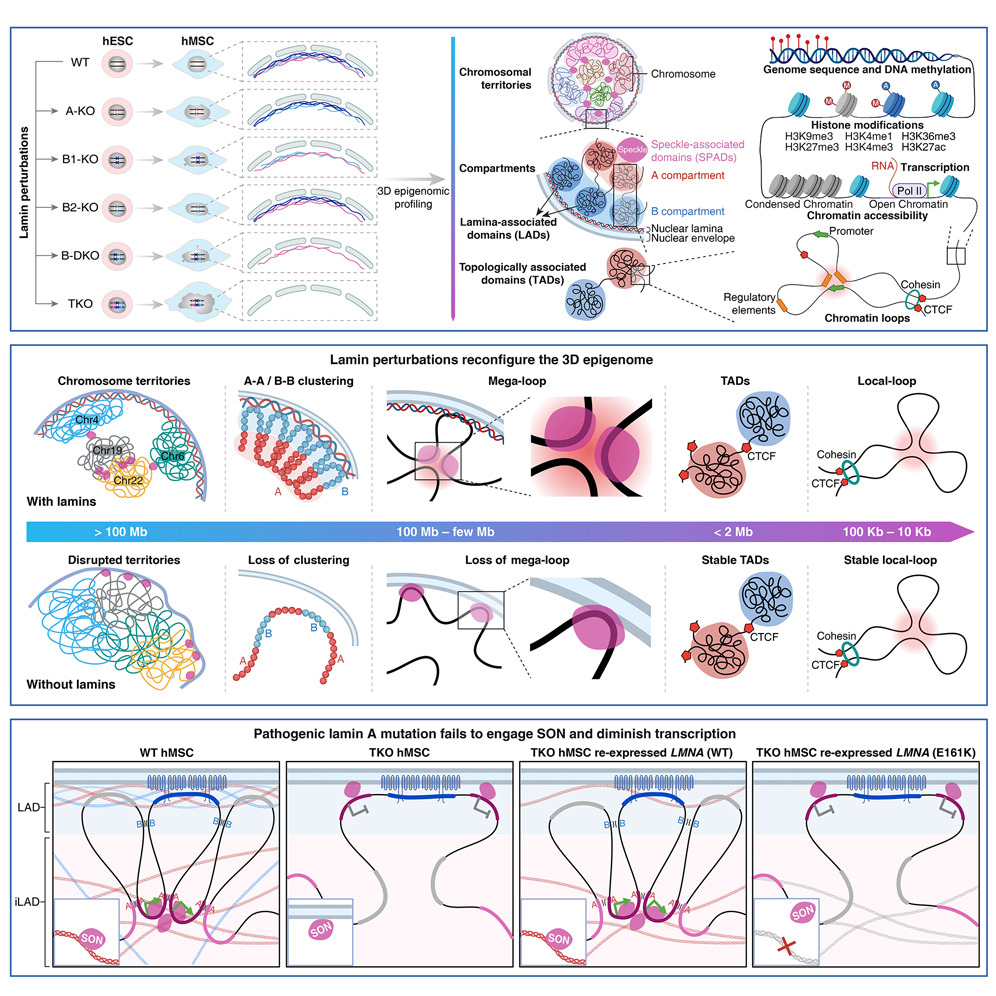
Great article about Lamins, proteins that help repair DNA breaks & stabilize chromosomes, preventing aging caused by epigenetic drift
When Lamins are missing, vital chromosomal loop structures that regulate genes unravel & aging accelerates
https://t.co/VY0esatTaD
Longevity NEWS: Anti-aging biotech @newlimit brings in @EliLillyandCo as investor, raising $45M, coming 5 months after the $130 million Series B round.
It was driven by new preclinical results that accelerated the timeline for starting IND-enabling studes.
https://t.co/jHQjURVdI3
Or a similar technology recently published by @altos_labs using the promoter of a senescence-related gene Cdkn2a (p16), to specifically express reprogramming factors OSK in senescent cells:
https://t.co/fazsQ5AHJz
New work by @RetroBio_@OpenAI advances the iPSC field by using AI to make far more efficient variants of the natural Yamanaka factors, overcoming a significant obstacle in reprogramming cells to pluripotency:
https://t.co/TqVYtUofbi
These new factors may also contribute to in vivo age reversal by reprogramming; however, controlling these powerful factors in vivo remains a challenge. They may be paired with other technologies, such as @DrMichaelWest developmentally-regulated iTR: https://t.co/RcfkP87LKj
On September 28th, I decided to stop rapamycin, ending almost 5 years of experimentation with this molecule for its longevity potential.
I have tested various rapamycin protocols including weekly (5, 6, and 10 mg dose schedules), biweekly (13 mg) and alternating weekly (6/13mg) to optimize rejuvenation and limit side effects.
Despite the immense potential from pre-clinical trials, my team and I came to the conclusion that the benefits of lifelong dosing of Rapamycin do not justify the hefty side-effects (intermittent skin/soft tissue infections, lipid abnormalities, glucose elevations, and increased resting heart rate). With no other underlying causes identified, we suspected Rapamycin, and since dosage adjustments had no effect, we decided to discontinue it entirely.
Preclinical and clinical research has indicated that prolonged rapamycin use can disrupt lipid metabolism and profiles [1], as well as induce insulin and glucose intolerance [2] as well as pancreatic Beta-cells toxicity [3]. Despite anecdotal evidence of rapamycin slowing down tumor growth, its effect in inhibiting natural killer cells [4] do raise concern for anti-cancer immune surveillance and cancer risk in the longer run.
Additionally, on October 27th, a new pre-print [5] indicated that Rapamycin was one of a handful of supposed longevity interventions to cause an increase/acceleration of aging in humans across 16 epigenetic aging clocks. This type of evaluation is the first of its kind, as most longevity interventions up to date have been tested against one or two aging clocks, leading to invisible biases and potential intended “cherry picking” of favorable clocks for the tested interventions.
Longevity research around these experimental compounds is constantly evolving, necessitating ongoing, close observation of the research and my biomarkers which my team and I do constantly.
Sources:
[1] https://t.co/clXah1mOuc
[2]https://t.co/mSlnpOYJRg.
[3]https://t.co/05ljueNWOM
[4]https://t.co/NIdYwzEilk.
Exciting! Belmonte lab reports OSK-reprogramming of old (p16+) cells increases wound healing & the lifespan of mice without causing cancer. Bodes well for human trials @lifebiosciences beginning next year 🚀🧵 https://t.co/MnAnl3mMBl
I heard from Michael West that Leonard Hayflick, who discovered cellular senescence (the Hayflick limit), passed away yesterday. He was the founder of the field and will be missed.
Here is a great interview of Dr. Hayflick by Dr. West.
https://t.co/0iPPSW7y7R
New preprint from the lab. Comparative analysis of previously established and novel reprogrammable mouse strains. Goal: generate new tools to achieve significant organismal rejuvenation and lifespan extension in wild type mice!
https://t.co/aL44z4TAS3
🚀 “Each month, we grow increasingly confident that age is reversible.”
Partial reprogramming holds incredible rejuvenating potential that could revolutionize medicine as we know it.
Great article by the CEO of @LifeBiosciences about the first clinical trial to test age reversal by cell reprogramming in humans 👏🏻 https://t.co/8YqgE6AXoq





















![GerontologyMike's tweet photo. In this, the first of 10 posts on age reversal by genomic reprogramming, I begin where the story really needs to begin; namely, the insights of the German naturalist August Weismann. Weismann proposed the term “immortality” to describe the unceasing lineage of cells called the “germ line” that perpetuates the species (that is sperm and egg that make babies that make sperm and egg that make babies, ad infinitum…). Weismann theorized that cells that make up the body (somatic cells) lost that capacity for immortal regeneration, a phenomenon I call “somatic restriction.” He stated “Death takes place because a worn-out tissue cannot forever renew itself, and because a capacity for increase by means of cell division is not everlasting, but finite [in the soma].” If you think about it, this means that the cells that we are made of (trillions of them) have no dead ancestors. Tracing them back in time, the ancestors of the cells that make us “us” have been proliferating since the first dawn of life on Earth, some billions of years ago. They will abruptly die in a few decades in us. The question we began to ask nearly a century after Weismann was, “can we transfer the capacity of immortal renewal characteristic of the germ line cells into our body, into the soma.” In the next post I will describe the first of such steps that led to the modern technology of immortality transfer.](https://pbs.twimg.com/media/HKeBkOmaoAAFiSA.jpg)